Å vokte kromosomer i mitosekorralen
Av David Coppedge, 8. juli 2024. Oversatt herfra
 Hodet mitt svømmer etter å ha lest flere nylige artikler om celledeling. Mangfoldet av involverte molekyler, og hvordan de kommer dit de trenger å være og gjøre det de trenger å gjøre - det er så forbløffende, det tigger beskrivelse. Jeg skulle ønske det var bedre måter å kommunisere til legfolk den emosjonelle effekten av å lære om cellebiologi, uten å drukne dem i sjargong som dette utdraget fra en artikkel om sentrosomer:
Hodet mitt svømmer etter å ha lest flere nylige artikler om celledeling. Mangfoldet av involverte molekyler, og hvordan de kommer dit de trenger å være og gjøre det de trenger å gjøre - det er så forbløffende, det tigger beskrivelse. Jeg skulle ønske det var bedre måter å kommunisere til legfolk den emosjonelle effekten av å lære om cellebiologi, uten å drukne dem i sjargong som dette utdraget fra en artikkel om sentrosomer:
Under interfase danner CEP57 et kompleks med CEP63 og CEP152, og fungerer som regulatorer for sentrosom--modning. Imidlertid er det molekylære samspillet av CEP57 med disse essensielle stillasproteinene uklart. Her demonstrerer vi at CEP57 gjennomgår flytende væskefase-separasjon, drevet av tre kritiske domener (NTD, CTD og polybasisk LMN). In vitro CEP57 kondenserer katalyserer mikrotubul nukleering via LMN-motiv-mediert tubulin-konsentrasjon. I celler er LMN -motivet påkrevd for sentrosomal mikrotubul aster dannelse. Videre begrenser CEP63 CEP57 -montering, utvidelse og mikrotubul polymeriserings -aktivitet.
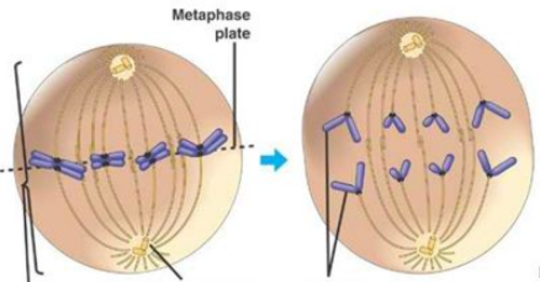
Bilde 1. Mitose -skjematisk fremstilling
Spesialister er komfortable med denne type snakk, men vi har en verden av studenter som blir fortalt at celler "dukket opp" ved en tilfeldighet, og "utviklet seg" til mennesker fra blinde, målløse prosesser gjennom millioner av år. Fakta over skriker "På ingen måte!" Men hvordan forteller vi ikke-forskere, uten å begrave dem i ukjente begreper? Analogier og bilder kan bare gå så langt. Jeg prøver mit beste, men enkle forklaringer kan ikke yte rettferdighet mot virkeligheten.
 For studenter hvis kunnskap om mitose kan være begrenset til lys-mikroskopbilder av delende celler, eller biologiske diagrammer på videregående skole av de fem stadiene i celledelingen (interfase, profase, metafase, anafase, telofase-"hm-hm, vil det være på prøven?' La meg prøve å pakke ut noen av de fantastiske underverkene som er gjemt i avsnittet ovenfor) Det kommer fra en artikkel i PNAS -lenke av 15 spesialister ved Institute of Bioinformatikk and Structurell Biologi i Taiwans National Tsing Hua University. Svar med applaus på hver linje:
For studenter hvis kunnskap om mitose kan være begrenset til lys-mikroskopbilder av delende celler, eller biologiske diagrammer på videregående skole av de fem stadiene i celledelingen (interfase, profase, metafase, anafase, telofase-"hm-hm, vil det være på prøven?' La meg prøve å pakke ut noen av de fantastiske underverkene som er gjemt i avsnittet ovenfor) Det kommer fra en artikkel i PNAS -lenke av 15 spesialister ved Institute of Bioinformatikk and Structurell Biologi i Taiwans National Tsing Hua University. Svar med applaus på hver linje:
*Noen molekyler går sammen for å kontrollere andre molekyler!
*Molekyler samles i avdelinger uten membran for å fungere i harmoni!
*Viktige ting vil ikke skje med mindre alle delene er sammen! - Og dårlige ting skjer hvis de mislykkes!
Bilde 2. Studenter blir ofte hjernevasket med materialisme
Biokjemi er ikke normal kjemi, der molekyler kolliderer som biler med støtfangere, og noen ganger slår seg sammen eller bryter opp. Dette er robot-fabrikkarbeid på et høyt nivå. Med unnskyldning til Jonathan MacLatchie, som kjenner sjargongen og skrev på et mer vitenskapelig nivå nylig (her), og til alle de andre praktiserende forskerne som snakker slik, i deres daglige rutine, må vi få høyet ned der kyrne kan spise det. Studentene blir så indoktrinert til materialistisk scientisme når de når høyere utdanning (hvis de søker en karriere på det nivået), at det blir vanskelig for dem å bryte med den evolusjonære konsensus, når de begynner å lære om de virkelige problemene i cellebiologi. Man må nesten oppnå ansettelse før man kan se uten evolusjons-filter. Det skjedde en dag for Michael Behe, da han stirret på en elektronmikrograf av en bakteriell flagellum, og funderte: "Det er en påhengsmotor. Det er ikke en sjansesamling av deler. " Hans tanke startet en revolusjon -lenke.
Med håp om å generere noe av ærefryktet jeg følte i lesingen min, kommer her oversettelsen av hva denne artikkelen avslørte. PHD -ene kan hoppe over lekpraten ved å klikke på lenken til forskningen.
Mitose -ranchen
 Da jeg prøvde å visualisere dette, tenkte jeg på følgende analogi. Si at en rancher må duplisere kyrne i en korral og skape to korraler med identiske antall og typer kyr. Han har først cowboyer plassere identiske kyr sammen i åk (åk representerer centromerer). De stiller kyrne opp mellom de to korralene, hode til hale. Så tau andre cowboys kyrne fra motsatte retninger, og kaster lasso på spesialiserte "horn" på åkene - Kinetochorer. Det tar noen forsøk for en cowboy å få lassoen på hornet på sin side av åket, men hver og en prøver å prøve til de lykkes. En inspektør sjekker at hvert åk-par har ett og bare ett tau på hver side, og at tauene er stramme. Når han gir et signal (sjekkpunktet), går en annen cowboy nedover linjen og bryter åkene. Cowboyene drar deretter kyrne inn i motsatte korraler, og en port lukkes mellom dem.
Da jeg prøvde å visualisere dette, tenkte jeg på følgende analogi. Si at en rancher må duplisere kyrne i en korral og skape to korraler med identiske antall og typer kyr. Han har først cowboyer plassere identiske kyr sammen i åk (åk representerer centromerer). De stiller kyrne opp mellom de to korralene, hode til hale. Så tau andre cowboys kyrne fra motsatte retninger, og kaster lasso på spesialiserte "horn" på åkene - Kinetochorer. Det tar noen forsøk for en cowboy å få lassoen på hornet på sin side av åket, men hver og en prøver å prøve til de lykkes. En inspektør sjekker at hvert åk-par har ett og bare ett tau på hver side, og at tauene er stramme. Når han gir et signal (sjekkpunktet), går en annen cowboy nedover linjen og bryter åkene. Cowboyene drar deretter kyrne inn i motsatte korraler, og en port lukkes mellom dem.
Realiteten i celledeling er mye, mye mer intrikat, men noe sånt skjer virkelig hver gang en eukaryot celle deler seg. Hundrevis av "cowboyer" kjenner rollene sine og vet når de skal gå i aksjon. I cellen er imidlertid "cowboyene" blinde og jobber i mørket. Begynner vi å bli overrasket ennå? Det er mye mer!
Bilde 3. Separering av kveg
Midlertidige møteplasser
PNAS -artikkelen avslører ny kunnskap om hvordan de rette molekylene kommer sammen i sentrosomet, en struktur som er kritisk for å trekke kromosomer fra hverandre under celledelingen. I mitose er det to sentrosomer, en på hver side av cellen. Sentrosomer er der den mitotiske spindelen sender ut "tauene" for å trekke kromosomene fra hverandre i datterkorralene. Innenfor hver centrosom vokser to fantastiske-symmetriske sentrioler vinkelrett på hverandre (se her). De vil være organiseringssentrene for spindelmikrotubulene. Mikrotubuler vil vokse ut fra sentrosomet og feste seg til kromosomene - en mikrotubul per søsterkromatid - ved spesialiserte koblingspunkter kalt Kinetochorer. Som et tau er hver mikrotubul sammensatt av flere strenger av tubulin, og gir stabilitet til hver spindelfiber.
Forresten, hvordan vet disse fjerne sentrosomene hvor mange kromosomer det vil være og komme med riktig antall mikrotubuler? Jeg stilte en KI -motor det spørsmålet. Den skrev:
 -Sentrosomet "vet ikke" antall kromosomer. I stedet blir prosessen med mikrotubuledannelse styrt av sjekkpunkter i cellesyklusen og regulatoriske proteiner. Under mitose festes hvert par kromosomer til en kinetokore, som deretter fester seg til spindelfibrene, dannet fra sentrosomet. Antall kromosomer bestemmer antall kinetokorer, som igjen hjelper til med å regulere antall mikrotubuler som trengs for riktig kromosomseparasjon. Det er en kompleks og koordinert prosess som involverer mange proteiner og reguleringsmekanismer.
-Sentrosomet "vet ikke" antall kromosomer. I stedet blir prosessen med mikrotubuledannelse styrt av sjekkpunkter i cellesyklusen og regulatoriske proteiner. Under mitose festes hvert par kromosomer til en kinetokore, som deretter fester seg til spindelfibrene, dannet fra sentrosomet. Antall kromosomer bestemmer antall kinetokorer, som igjen hjelper til med å regulere antall mikrotubuler som trengs for riktig kromosomseparasjon. Det er en kompleks og koordinert prosess som involverer mange proteiner og reguleringsmekanismer.
Bilde 4. Svar fra KI
Ok vel; Poenget oppfattet!
Tilbake til de "midlertidige møteområdene" ved sentrosomer. Sentrosomet har ingen membran. Men når CEP57 (sentrosomprotein nr. 57 av dusinvis kjent så langt) er til stede, dannes en midlertidig barriere rundt de nødvendige ingrediensene. Dette skjer med "væske-lipidfase-separasjon", noe likt hvordan det dannes oljedråper i vann (se min forrige artikkel om kondensater her https://evolutionnews.org/2021/12/caltech-finds-amazing-role-for-noncoding-dna/ ). Men først må det midlertidige møteområdet (Peri-Centriolar Matrise, eller PCM) vokse med en størrelsesorden, fra 300 nanometer til mikrometer, ettersom alle nødvendige ingredienser samles. "Menneskelig CEP57," sier de, "er et kveilespole stillas ved PeriCentriolar Matrise (PCM), som kontrollerer sentriolduplisering og sentrosommodning for trofast celledeling."
 "Før begynnelsen av mitose gjennomgår sentriolen sentriol-til-sentrosom-konvertering ved å rekruttere mer sentrosomale komponenter og utvide PCM til mikron-størrelse. Denne ekspansjonen fører til en økning i mikrotubul kjerne -faktorene, noe som letter den raske monteringen av den mitotiske spindelen under mitose. Det er et sentralt spørsmål om hvordan PCM samles i et tett rom beriket med hundrevis av forskjellige proteiner i det humane sentrosomet under PCM -ekspansjon. Væske-VæskeFaseSeparasjon (VVFS) er et overbevisende konsept for å belyse de organisatoriske prinsippene som ligger til grunn for membranfrie organeller. I VVFS-systemer driver multivalente interaksjoner gjennom foldede domener eller iboende uordnede regioner, og resulterer i dannelse av dynamiske biomolekylære kondensater, tilgjengelig for kognate klienter."
"Før begynnelsen av mitose gjennomgår sentriolen sentriol-til-sentrosom-konvertering ved å rekruttere mer sentrosomale komponenter og utvide PCM til mikron-størrelse. Denne ekspansjonen fører til en økning i mikrotubul kjerne -faktorene, noe som letter den raske monteringen av den mitotiske spindelen under mitose. Det er et sentralt spørsmål om hvordan PCM samles i et tett rom beriket med hundrevis av forskjellige proteiner i det humane sentrosomet under PCM -ekspansjon. Væske-VæskeFaseSeparasjon (VVFS) er et overbevisende konsept for å belyse de organisatoriske prinsippene som ligger til grunn for membranfrie organeller. I VVFS-systemer driver multivalente interaksjoner gjennom foldede domener eller iboende uordnede regioner, og resulterer i dannelse av dynamiske biomolekylære kondensater, tilgjengelig for kognate klienter."
Bilde 5. Centrosom -sentralt i celledeling (mitose)
"Hundrevis av ulike proteiner." Fikk du tak i det? Hvordan ender alle de rette i det membranløse kondensatet? Hvordan kunne blind evolusjon noen gang oppnå et slikt møte? Interessant nok spiller de "iboende uordnede regionene" av noen av disse proteinene, som kan ha blitt ansett som dårlig designet av noen, en nøkkelrolle i faseseparasjonen. Legg også merke til at kondensatet er "dynamisk" og "tilgjengelig" for de "kognate klientene" som hører hjemme der, samtidig som ikke-medlemmer holdes utenfor.
Forfatterne påpeker at en mangel på akkurat denne komponenten, CEP57, resulterer i uorganisering av PCM, og en forferdelig sykdom:
"CEP57-mutasjoner er genetisk knyttet til Mosaikk-Variegert Aneuploidi (MVA), en sjelden sykdom preget av et unormalt antall kromosomer. MVA -syndromet manifesterer seg ved forskjellige lidelser, inkludert skjelettanomalier, mikrocephaly og kreft i barndommen."
CEP57 er ikke den eneste viktige delen. "CEP57, CEP63, CEP152, CEP192, CDK5RAP2 (CEP215) og pericentrin er essensielle stillasproteiner for PCM -integritet."
Hva viser din ærefrykt-måler så langt?
 Det er vanskelig å formidle undringen fra disse realitetene uten å bli overdynget av sjargong og detaljer. For mer ærefrykt, legg til disse betraktningene:
Det er vanskelig å formidle undringen fra disse realitetene uten å bli overdynget av sjargong og detaljer. For mer ærefrykt, legg til disse betraktningene:
*Alt dette foregår i rom for bittesmå til å se med det blotte øye.
*Forskere har oppdaget de fleste av disse intrikate detaljene i løpet av levetiden til mange av dem i dag.
*Sekvensen av aminosyrer i hvert involvert protein er altfor usannsynlig til å ha sin opprinnelse i tilfeldighet.
*Flere millioner celler deler seg hvert sekund i kroppene våre.
*Celler har delt seg siden begynnelsen av livet på jorden.
*Nøyaktigheten av celledeling er så ekstraordinært høy, mange dyr i dag er gjenkjennelige fra sine kolleger i fossilregisteret.
Bilde 6. Ny vinkling på biologi -systembiologi
Vi er virkelig privilegerte ved å se detaljer om underverker som var skjult for menneskers øyne i tusenvis av år. Hvis romere og babylonere og eldgamle kinesere ble imponert over synet av en baby ved fødselen, hvor mye mer burde vi være lamslått, stumme, virkelig ærbødige om hva biokjemikere lærer i dag om realiteter for små for menneskelige øyne? Visst, noen observasjoner som ondskap og lidelse er vanskelig å forstå, men selv disse er bedre tilgjengelige for forklaring i designsammenheng -lenke. Som min college -biologi professor pleide å si: "Det fantastiske er ikke at vi blir syke. Det fantastiske er at vi noen gang er friske, "med tanke på hvor mange ting som må fungere riktig hvert øyeblikk, hver dag. Bli aldri selvtilfreds med disse realitetene som finner sted i oss. Vi er vitne til intelligent design på et nivå som aldri er forstått tidligere, gjennom hele menneskehetens historie.

Om forfatteren: DAVID COPPEDGE -kredit til David Coppedge (Bilde 7)
David Coppedge er en frilans vitenskapsreporter i Sør-California. Han har vært styremedlem i Illustra Media siden grunnleggelsen og fungerer som deres vitenskapskonsulent. Han jobbet ved NASAs Jet Propulsion Laboratory (JPL) i 14 år, på Cassini-oppdraget til Saturn, til han ble kastet ut i 2011 for å dele materiale om intelligent design, en diskriminerende handling som førte til en nasjonalt publisert rettssak i 2012. Discovery Institute støttet saken hans, men en ensom dommer dømte ham mot ham, uten forklaring. En naturfotograf, friluftsmann og musiker, David har B.S. grader i realfagsutdanning og i fysikk og holder presentasjoner om ID og andre vitenskapelige emner.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund